Previously we discussed the transition from acute to chronic stress responsiveness, and how this transition might explain the excess anxiety seen in post-traumatic stress disorder (PTSD) sufferers as a consequence of a recruitment of amygdalar CRF (a vital brain juice involved in both panic and depression). How might we explain the hyperarousal and hypervigilance that is also associated with PTSD?
The Locus Coeruleus and Attentional Arousal
Arousal and stress have long been associated increased activity in brain catecholamines, such as adrenaline and noradrenaline. There are two major bundles of nor-adrenaline-containing neurons ascending from the brainstem to higher structures, commonly referred to as the ventral and dorsal noradrenergic bundles. The ventral bundle projects to subcortical structures, including the motor neurons in the hypothalamus that drive the neuroendocrine stress response. Putting noradrenaline directly on to these motorneurons produces an HPA axis stress response. This lower bundle is known to mediate many "bottom-up" stress responses, such as responses to hunger and pain.
The higher, dorsal bundle, originates in two tiny, bilateral nuclei in the pontine brainstem known as the locus coeruleus ("blue spot"), which supplies virtually the entire cortex with its sole supply of noradrenaline. The locus coeruleus is highly responsive to arousing or salient stimuli, and when activated it dumps noradrenaline into places like the neocortex and hippocampus to alter their electrical activity and information processing abilities. In sensory cortex, for example, LC activity can increase the throughput of sensory signals. LC also projects strongly to areas involved in attentional processes, such as the superior colliculus. Say you are watching television and in the very, very corner of your eye you see something, like a mouse, move against the baseboard, and you suddenly turn toward it, but it’s gone. "Holy crapola, Motorola! I swear I just saw a mouse!" That is your superior colliculus at work, being attentive to the environment around you, when "you" and your focus of consciousness are paying attention to other things. Just about any stimulus that arouses behavioral responses, such as orienting responses, also excites the LC. During hyperarousal, you tend to see more things in the corner of your eye.
Yerkes-Dodson Law: Performance as a Function of Arousal
It has long been known that performance on a wide variety of tasks is a function of arousal. At very low levels of arousal, an organism is non-alert and inattentive, and in the limit either asleep or dead. Performance under low arousal is extremely poor. Conversely, when organisms are extremely aroused they tend to be highly distractible and unable to focus on relevant tasks, and again performance suffers. Where performance peaks is at intermediate levels of arousal, when the organism is attentive, but not distractible. This is a very consistent finding, and the inverted-U shaped function looks like this:
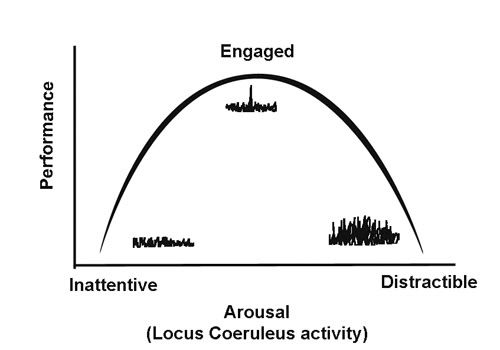
Optimizing Tasks: LC Activity Modes and Utility
The the squiggles in the illustration above also show different modes of neuronal activity in LC. The LC has two modes of firing, a baseline mode of tonic random firing, and a phasic mode that becomes locked to relevant sensory events and motor acts related to focal attentional tasks. At very low and very high levels of arousal, the tonic mode predominates. Whereas at peak performance, the LC delivers phasic burst. According to Aston-Jones and Cohen (2005):
In a phasic mode, bursts of LC activity are observed in association with the outcome of task-related decision processes and are closely coupled with behavioral responses that are generally highly accurate. In a tonic mode, LC baseline activity is elevated but phasic bursts of activity are absent, and behavior is more distractible.
The LC is Used to Focus Attention on Relevant Events & Tasks.
When the utility of action wanes, the LC, instructed by specific areas of cortex, can withdraw support for different tasks. So what areas of cortex are instructing the LC about giving or withdrawing support for various tasks? One is the orbital-frontal cortex, an area of the brain that is involved in storing and comparing value-laden and emotional memories. Another is the anterior cingulate cortex, which is involved in conflict processing. Together these two areas of cortex can instruct the LC on (a) whether external events are important, (b) which are more important, (c) what response one should select given the potential for making several competing responses.
According to Aston-Jones and Cohen:
We propose that within the context of a given task, phasic activity of the LC-NE system facilitates behavioral responses to the outcome of task-specific decision processes, filtering responses to irrelevant events. By selectively facilitating responses to task-relevant processes, the LC-NE phasic response serves to optimize the trade-off between system complexity (which can support a broad range of functions) and efficiency of function (optimizing performance in the current task). We further propose that the LC-NE system is responsive to ongoing evaluations of task utility (that is, the costs and benefits associated with performance), provided by input from frontal structures.
In short, in normal fucntioning, LC activity focuses cortical processing on relevant tasks. Below are some examples of LC firing in response to various stimuli (a cartoon adapted from Aston-Jones & Cohen, 2005). In this task, a monkey is trained to fixate on a spot at the center of a sceen, and bar press for juice reward only when the appropriate target stimulus arrives. Non-target (irrelevant) stimuli are also presented.
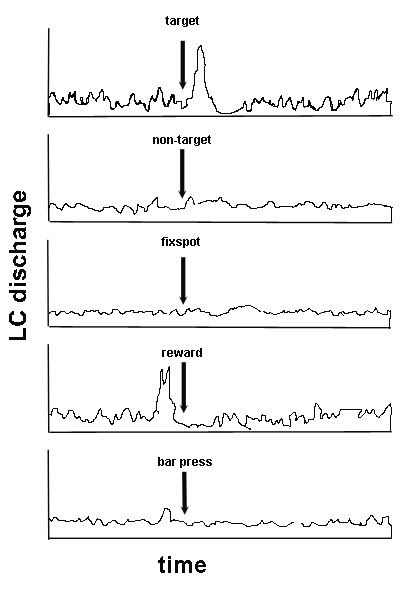
The horizontal axis shows time (about 2 sec total), and the vertical deflections show LC firing. Arrows show when various events occur. The panels are out of order, but you'll get the picture anyway. Before the target stimulus arrives, LC firing is tonic, whereas right after the target arrives LC fires a phasic burst to focus the appropriate cortical circuit. The arrival of the irrelevant non-target stimulus does not change the firing rate. Nor does the arrival of the fixation spot. These stimuli also have no value. Now, how in the world does LC firing become phasic just before the arrival juice reward? That burst corresponds to the "click" of the solenoid that delivers the juice. A relevant click. Finally, there is minor burst in LC phasic firing just before the monkey presses the bar, another relevant event.
So what happens when reward disappears?
When utility persistently wanes, changes in LC-NE tonic activity withdraw support for task performance, facilitating other forms of behaviors that serve to explore alternative sources of reward. These functions are accomplished by the neuromodulatory effects of NE release at cortical target sites, modulating the gain (responsivity) of processing in cortical circuits responsible for task performance. The different modes of LC activity adaptively adjust the gain of these cortical circuits, both phasically and tonically, facilitating or disengaging task-specific responses.
Via instructions from value-laden and conflict processing areas of cortex that monitor ongoing events, LC withdraws support for formerly relevant tasks by reverting to the tonic firing mode that more generally samples incoming information, generating complexity at the expense of task-specific focus. An increase in tonic firing "serves to explore alternative sources of reward."
Now, it is my interpretation that when Aston-Jones & Cohen say that tonic activity "serves to explore alternative sources of reward," they don’t mean reward as only good things. They just happened to be on the topic of food reward at the time. I will bet my bottom dollar they more generally meant any kind of significant event whether pleasurable or aversive, because the LC should be able to direct adaptive, focal phasic responses to any significant event, not just the pleasureable ones. If you need to defend yourself, you better be able to focus your attention and the appropriate stimuli and your responses on the appropriate reactions. In fact, if that solenoid click that signals juice delivery focuses arousal, you can bet that the magazine click of a gun will do the same if you're on night patrol outside the green zone. If there is no appropriate focal stimulus to focus on in order to maximize your utility, then you disengage and begin scanning the environment for that click using the tonic activation mode.
Without going into detail, increases in tonic activation are also associated with greater "false alarms" on signal detection tasks. Sort of like that mouse running along the baseboard that you may or may not have seen.
The Effects of CRF on LC Activation:
In a previous we talked about how chronically elevated stress steroids act to upregulate and recruit central stress networks involving amygdalar CRF. What I didn’t say was that stress steroids upregulate CRF in anatomically related areas, such as the bed nucleus of the stria terminalis, as well. This is important because these CRF-containing, anxiety-mediating nuclei project heavily to the core and surround of locus coeruleus. Now, look at what CRF does to activity in the locus coeruleus (cartoons adapted from Rita Valentino's lab; Curtis et al 1997).
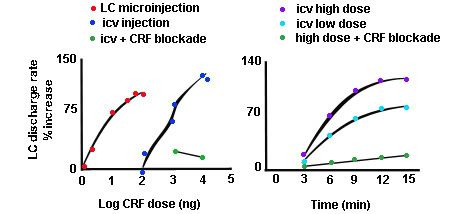
The left panel shows that LC discharge rates dose-dependently increase with increasing doses of CRF. The red dots show discharge when CRF is microinjected directly into LC, whereas the blue dots show discharge rates when CRF is put into the brain ventricles. Obviously, lower doses of CRF are required when put directly into LC. The green dots show that activation of LC is blocked when CRF is given in conjunction with a CRF receptor blockade, demonstrating CRF specificity.
The right panel shows the evolution of LC discharge rates over time with higher (blue) and lower (aqua) doses of CRF injected into the ventricles. The discharge rates increase over time, are dose-dependent, and are again blocked by CRF receptor blockade.
What this says is that chronically elevating CRF transmission as a result of chronic exposure to elevated steroids is going to push LC discharges from some optimal level discharge where phasic firing enhances performance into a state of hyperarousal where tonic firing swamps out the ability to respond to task relevant events.
Excerpt from Ilona's excellent book on PTSD:
Once he got home, however, Dwyer found himself feeling detached and fidgety, and struggled to perform the common, everyday tasks that we all take for granted. Once an extrovert, he now hated being out in noisy crowds. At restaurants, he always kept his eye on the door...
Dwyer's older sister said of her brother, "To me, he hasn't fully left Iraq"
I have to say, Ilona's book is so information-rich, that when I was reading it, it was like "ding, ding, ding!" from page to page.
Now there are at least two ways to interpret increased distractibility and hypervigilance due to stress steroid--> CRF--> LC facilitations with respect to PTSD.
- The value-laden cortical system that instructs LC has increased the relative value or salience of war (or other traumas for that matter) compared to normal life, and continually instruct the LC to continue scanning for something really important, like signs of further trauma (nothing in normal life is important anymore compared to trauma).
- The sheer hyperarousal of LC drowns out signal relative to noise, and the PTSD is simply distractable. Filtering and focus are impossible due to the high background noise.
I suspect both are true, but I don’t know of any data on this. We will get to why I think number one is important in a later diary on value-laden memory.
