I've been struggling a bit with the next chapter in my series and part of the reason is that it is difficult to discuss evolutionary ideas without talking about inheritance and going into long digressions about the nature of genetics. So I thought I would devote a diary to discussing genetics and inheritance. I could then refer to this diary in later diaries. I also thought it might help kossacks who are not highly knowledgeable about genetics think about some of these issues in their personal and political lives. I'm going to use a fairly non-technical approach in my explanation. This is deliberate because IMHO in trying to understand inheritance it is easy to get bogged down in the details of molecular biology and lose sight of the big picture.
As mentioned in my previous diary the prevailing theory of inheritance in the 19th century was blending inheritance in which the characteristics of both parents were blended in the offspring. It is easy to understand the appeal of this concept - it makes intuitive sense and many of our observations appear to bear it out.
Mendel's View of Inheritance Gregor Mendel's experiments demonstrated that inheritance was actually particulate. In other words there were units of inheritance that were passed to offspring from both parents and that these units (which today we call genes) remained distinct from each other even though their effects (the characteristics of the organism or phenotype) might be blended.
Mendel's work was published in the 1860s to a complete lack of recognition. It was rediscovered independently by multiple scientists in 1900 and completely revolutionized biology. Fifty years later the structure of DNA was determined and the field of molecular genetics really got going. It was recognized that genes are sequences of DNA which in turn code for sequences of RNA which code for amino acid sequences. Those of you with any background in molecular biology will recognize that this is a gross simplification. What I am trying to do is not give a history of genetics but rather to illustrate the fundamental way in which genetics is viewed and taught.
Millions upon millions of children and undergraduates are taught about genetics in a way that I think can lead to confusion about how genes operate in most cases. This isn't due to any ideological bias but rather that the examples used to illustrate the basic principles of genetics are atypical. They are used because their atypical nature makes the underlying process of inheritance more obvious than is the usual case.
What the heck am I talking about? Mendel is widely theorized to have 'doctored his results'. Not in the sense that he made up data or changed results but that he was almost certainly quite selective in only publishing results that supported his hypotheses.
Mendel proposed the unit of inheritance (genes) existed in pairs within individuals. Each member of a pair is called an allele. Individuals could thus have two identical alleles and be homozygous (identical sequences of DNA) or two different alleles and be heterozygous (similar but not identical sequences of DNA). Each individual passed one of the alleles for each gene on to each of its offspring.
In order for Mendel to easily detect this pattern in his crosses of pea plants he had to pick traits of the plants that were inherited in particular ways. First, and most importantly, he only used traits where the variation in his populations was affected by a single gene (biologists would use the term locus (plural loci) here but I will use gene to mean the same thing as locus). Secondly each gene had two different alleles which led to two very distinct forms the trait (two different phenotypes). Third the two different alleles had a clear relationship to one another such that in a heterozygote the phenotype was the same as one of the homozygotes but not the other. This is known as complete dominance.
Here is an example from Mendel. Flower color in peas was controlled in his population by a gene with two alleles called P and p.
Genotype Phenotype
PP Purple Flower
Pp Purple Flower
pp White Flower
It is fairly straightforward to detect the pattern of inheritance in this kind of system by doing crosses or by following family histories. Human geneticists were able to figure out the pattern of inheritance for a number of genetic diseases (hemophilia, Huntington's disease, cystic fibrosis, etc.) because they fit this pattern. The phenotype fell into two well defined categories (had the disease or didn't have the disease) and a single gene with complete dominance.
(simple) Mendelism is deterministic genetics
We can write out (part of) Mendel's concept of inheritance as an equation (this equation is symbolic - not a real equation).
genotype = phenotype
What this means is that in the examples we've discussed the particular set of alleles you have for a gene completely determines your phenotype
This can be broken down into the following
alleles + interaction between alleles = phenotype
What I mean by this is that it isn't just the alleles you have but the interaction between them that determines the phenotype. The p allele has a different affect on the phenotype in an individual that is Pp than it does in an individual that is pp.
Complications The problem is that most characteristics (phenotypes) are not inherited in that way. These cases lead to a simplistic and overly deterministic view of how genes operate in most cases. Generally the traits we are most interested in are quantitative/complex traits. These kinds of traits don't have two possible phenotypes but a continuous range of phenotypes. They are influenced by many genes and by the environment.
There are two additional factors that need to be considered in inheritance. These are interactions between different genes. This is known as epistasis and is something that gives students headaches. The second is the effect of the environment on the trait.
Epistasis - Interactions between Genes - An Example Seeing as this is Daily Kos we will use cats to demonstrate epistasis. I'm doing so with a bit of a reservation as we are still dealing with very simple patterns of inheritance for illustrative purposes which is the very thing I want to get away from.
One gene that influences cat fur color is called agouti. It has two alleles A and a
In cats that have genotypes AA or Aa the individual hairs have bands of different colors giving the fur a grizzled appearance. In cats that have genotype aa each hair is a solid single color (most commonly black).
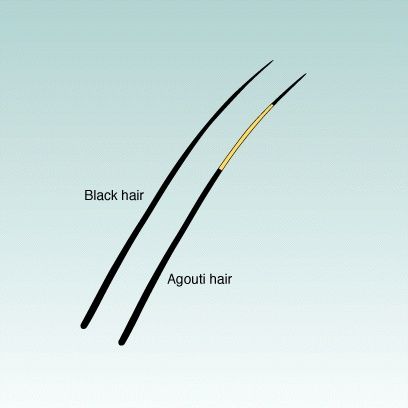
Agouti and Black Hairs
There is a second gene called tabby. It has three alleles. The T allele produces the common tabby pattern with parallel stripes of black fur (solid colored hairs) with agouti fur (banded hairs) between the black stripes. The Ta allele produces the abyssinian color pattern in which all the fur is agouti. The t allele produces what is known as 'classic tabby' which is seen in show cats but, at least in my experience, rarely in the general cat population. The classic tabby has a more complex pattern of black and agouti fur with swirls rather than fairly straight lines. The t allele is recessive to the other two. T and Ta have what is known as incomplete dominance. An individual with one of each (e.g. TTa) usually has black striping on the face, legs, and tail but not on the body which is all agouti.

A classic tabby pattern
The tabby gene only affects coloration on cats with at least one A allele (e.g. AA or Aa). In aa cats it will have no effect whatsoever. This is an example of epistasis.

The black cat could have the same tabby genotype as its basket mate the mackerel tabby
So I'm going to expand my equation to say
alleles (for many genes) + interaction between alleles of one gene + interaction between alleles from different genes = phenotype.
First Take Home Message on Genes and Inheritance The effect of an allele on the phenotype depends not just on the allele but on the genetic background, the particular combination of alleles across multiple genes in an individual.
So we have muddied the water of inheritance quite a bit and we haven't even got to the environment yet.
The Effect of the Environment Let's stick with cats for a minute. The Siamese pattern of coat coloration is caused by an allele of a particular gene (a different allele will produce a solid white cat). This allele affects the deposition of pigment in hairs based on temperature. Pigment is deposited in fur on regions of the body that are slightly lower in temperature (ears, face, ends of legs, end of tail). Pigment is not deposited in the torso. This can be tested by artificially cooling (e.g. with a cold pack) a part of the torso long enough to grow new fur or by warming the feet long enough to grow new fur.

This shows two things. First is that the environment (temperature in this case) can also influence the phenotype and secondly that the effect of the environment may vary with genoytpe. The fur color of non-siamese cats is not affected by temperature.
So we end up with this as our equation
alleles (from many genes) + interaction between alleles of one gene + interaction of alleles from different genes + environment + interaction between alleles and the environment = phenotype.
OK so I may have convinced you that genetics is complicated - so what? The second take home message is that the notion of nature vs nurture is a false dichotomy. There is input from both and often in complex ways.
Variation and Quantitative Genetics Now lets rephrase this in terms of variation. It isn't really practical to look at a trait in an individual (for example my height) and ask what proportion of my height is due to my alleles, what proportion is due to epistasis, and so on. My height is the result of developmental processes that are now done and even if they weren't done, measuring the input of each factor on the phenotype an individual is not really possible. How could you really quantify how much of your height was due to the environment and how much is due to genes? What would you actually measure? This is why I said that the equations above are symbolic.
Quantitative geneticists, biologists who study the inheritance of traits influenced by many genes, have solved this problem by by measuring how variation in these factors across individuals influence variation in the phenotype in the population.
In non-human organisms this is done using breeding experiments. In humans cruder estimates are possible in a variety of way such as twin studies.
So my equation would be rewritten as a real equation.
VA + VD + VI + VE + VGE = VP
VP is the total amount of variation in the trait in question (this is measured using a metric called the variance - if you are not familiar with it don't worry about it). In my example it would be the amount of variation in height in the population to which I belong. If everyone in the population was exactly the same height then the variation is zero. VA, VD, VI, VE, VGE are the amount of the variation due to alleles, interactions among alleles of a gene, interaction of alleles from different genes, the environment, and the interaction between genes and the environment. The experiments done by quantitative geneticists partition the total variation in the trait into these different components.
The ratio VA/VP is known as the heritability. This is the proportion of the variation in the trait that is due to alleles independent of genetic background. The heritability can vary between 0 and 1. A heritability of 0 means that parents do not resemble their offspring for this trait any more than random members of the population. A heritability of 1 means that the phenotypes of the parents will exactly predict the phenotypes of their offspring.
Heritability has been used as a metric to look at the inheritance of many traits. Because it is a ratio it can be used to compare very different types of traits. However it can be very easily misused.
Problems and Misinterpretations of Heritability
Problem 1. Heritability is a measure of a trait in a particular population. It depends on the values of a number of different factors which are likely to vary in space and time. So a measure in one population might not be as informative in a general sense as you might think. Also it is a ratio based on data on variation WITHIN a population. It doesn't tell you anything about variation among populations.
To illustrate this here is a simplistic example. Imagine we have a population of humans that set out from Europe on two boats in 1500. Both boats had populations with variation in skin color but the same average skin color and same amount of variation. The two boats are separated at sea and one ends up in Newfoundland and the other ends up in Trinidad. If an alien arrived on earth ten years later and took samples of humans from both populations and did breeding experiments the alien might well find that WITHIN each population there was genetically based variation for skin color. The alien might also note that the average skin color differed BETWEEN the two populations. In this case the variation BETWEEN populations would be wholly environmental due to differing exposure to solar radiation.
Problem 2. Heritability is a ratio and its magnitude depends on two different factors. The variation in number of eyes in human populations is essentially zero (everyone has two eyes) and thus the heritability is also zero. No one believes that the number of eyes we have is not influenced by genes.
Less trivially there have been quite a few reviews of heritabilities of different kinds of traits in animals. Anatomical traits such as the length of a certain bone or height tend to have high heritabilities in 0.7-0.9 range. Behavioral traits or what are known as life history traits (fertility, growth rate, lifespan) tend to have lower heritabilities in the 0.1-0.5 range. There are a couple of things to note about this. First, most of these estimates come from laboratory studies where the environmental variation is minimized and thus lower than it is in a natural population (thus increasing the heritability). Secondly it turns out that in these cases heritability is lower not because VA is smaller but because VE is larger resulting in a larger VP. This might seem like a trivial distinction. What is saying is that a trait has a lower heritability not because it is not influenced by genes but because it ALSO is very highly influenced by the environment.
Problem 3. Accurately estimating the heritability of a trait (or other quantitative genetic parameters) is very difficult and time consuming and is often done imperfectly in non human studies and very frequently if not universally done imperfectly in human studies. As you might imagine, teasing apart the different components in our equation in the real world is very difficult. One persistent type of problem is maternal effects. Nongenetic influences from the mother (the amount and types of nutrients laid down in the egg, the biochemistry of the uterine environment, etc.). Maternal effects will cause siblings to resemble one another but are not based on genetics.
Separating these types of subtle and deep-rooted environmental effects from genetic effects requires breeding large numbers of individuals of know relatedness to one another in a laboratory environment in which, to the extent possible, the effect of the environment is not correlated with family membership.
Human quantitative genetics studies inherently lack the power of these laboratory studies. Modern molecular techniques such as QTL analysis are greatly improving studies of the genetics of complex traits in humans. However the problem remains to some extent - you can't control either the environment or the extent of relatedness and so human researchers always have much less power.
I'm certainly not implying that it useless to study the genetics of complex traits in humans or that all such studies are bad. However they should be viewed in light of the formidable logistical obstacles that they face. Press releases often talk about finding a 'gene for schizophrenia' or a 'gene for alcoholism'. The simple model of gene action I started with can give you a very different notion of what that means relative to the more complex model we used at the end. The 'gene for alcoholism' may be nudging people slightly in that direction and possibly only in people with certain other genes or in certain environments.
Postscript: Scientific Problems With Eugenics
Eugenics was a powerful movement in the late 19th and early 20th centuries. The central concept was to use the principles of artificial selection (as used on domesticated animals and plants) to improve human populations. Nazism ended eugenics as a popular ideology although it does crop up regularly. I'm not going to discuss the ethical and other social problems in eugenics, most of them are pretty obvious. I will note that prior to Nazism eugenics was espoused by figures across the political spectrum and even by some very notable evolutionary biologists. I'm going to restrict my comments to the reasons why eugenics is bad science.
1. Populations and individuals. A major problem is a failure to think clearly about comparisons of individuals within a population and comparisons of different populations. If you are interested in IQ for example if is very clear that the rich and the poor would have to be considered different populations as the environment in which children of each group develop is so different.
2. Measurement. If you are looking to improve a characteristic it has to be measurable in a consistent manner. Not a big issue if you are measuring height but the research on intelligence is riddled with the difficulty of measuring a rather abstract quality (intelligence) in a general, unbiased manner.
3. Relative input of genes and environment. Most of the traits of interest to eugeneticists were things like intelligence, health, fertility, etc. These are all characteristics that likely have a very high environmental input. Given the extremely non-random nature of our social environment, artificial selection would likely be relatively ineffective in 'improving' traits relative to improving the environment.
The following example illustrates all three of these problems. Imagine a situation in which a pig farm wishes to improve the protein content of bacon produced by its pigs. Each pig family is fed either low protein food or high protein food. Not surprisingly the pigs fed higher protein food have higher protein bacon. The farmer wishes to perform to artificial selection to achieve the goal of overall improved protein content.
The following problems with an artificial selection plan become apparent.
Two populations (based on food input) are mixed together in the analysis
An environmental factor (food type) likely to highly impact the phenotype of interest is non-randomly distributed across familites
The environment is having a larger impact than genes.
How should the farmer increase protein content? In this case the logical thing to do would be to feed all the pigs the high protein food. In other words change the environment rather than the genes.
It is easy (I hope) to see this rather poorly-managed pig farm as a metaphor for human society. In addition to its moral short comings eugenics was not a practical methodology for improving human characteristics. Improving the environmental factors is both more moral and more effective.
