For those of you who are gluttons for punishment here is the third diary on color and biodiversity.
Part I: Introduction to Color Basics and Color Genetics
Part II: Development of Color Patterns, Color as an Antipredator Mechanism
This time we're going to talk about animals that have a great deal of control over their body color and about how the perception of color may vary.
A slight change of schedule for the rest of this series.
Next Sunday - no diary
Jan 2 - Color and plant/animal coevolution
Jan 9 - Color and mate choice
Jan 16 - Color variation within a population
It is possible that I may get busy after the semester starts and have to skip a week. But the last two topics are ones I about which I am well versed and should be easy to write.

Squirrel treefrog on car.
I. Color Genetics and Development and Model Systems.
First up a few comments/additions to the discussion of the developmental control of color in animals. Explaining developmental biology is not the easiest thing in the world. I'll write a more detailed diary about it at a later date. What I'd like to do right now is talk a bit about model systems and understanding biological diversity.
A lot of biology is done on a relatively small amount of species. Lab mice, Drosophila (fruit flies), yeast and a few other organisms are the subjects of a vast amount of basic lab research. Even ecology has species and habitats that have been the subject of a disproportionate amount of research.
All model system images from Wikimedia Commons

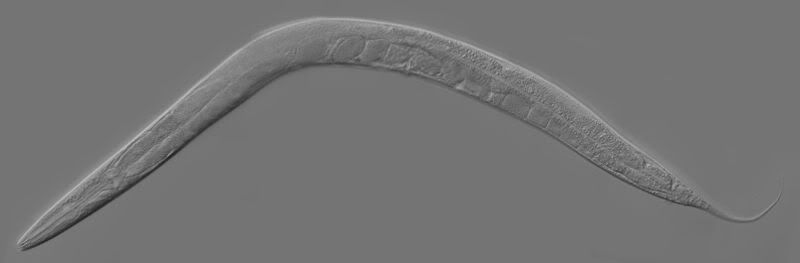
Drosophila melanogaster and Caenorhabditis elegans The former is the classic model organism for genetics and molecular biology and is used in other fields as well. The latter is a model for molecular studies of development because it has a fixed number of cells as an adult.

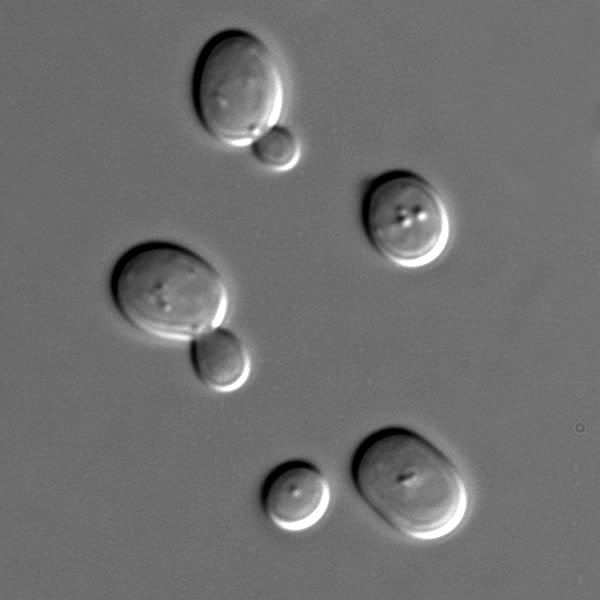
Zebra fish and yeast. Zebra fish have transparent embryos and are models for studying vertebrate development. Yeast are single celled but have cells similar to our own compared to bacteria. They are models for genetics and cell/molecular biology.
These are known as model organisms or model systems. The idea is that they are models for all (or some large subset) of the other organisms, communities etc. out there. By studying them you hopefully learn things that can be generalized across large chunks of the diversity of life.
Why proceed this way? Several reasons. One is that the diversity of life is immense and there is no way to study everything in every species and system. Secondly, the model species are usually chosen because they have properties that make them tractable for addressing a particular problem. For example fruit flies are excellent model organisms for the study of genetics. They have a rapid life cycle, can be reared in large numbers in a fairly small space with little maintenance. However they are a terrible modern organism for ecology. They are too small to individually mark and they are highly mobile so most individuals would never be found again after an initial capture.
Thirdly once a particular species/system has been studied in detail then the findings of those studies allow more sophisticated questions to be asked. So rather than starting back at square one with a new species/system it makes sense to carry on with the one you have.
Ecological, behavioral, and evolutionary biology also have model systems. Because the questions asked are often more specific there are many more model systems.
The rocky intertidal zone is commonly used to study how predator prey relationships shape communities. It has the advantage that most of the species involved including the top predators such as the -Pisaster_ seastar in this picture are easily found and not highly mobile so they can be easily censused.

Crickets are common study organisms for a lot of behavioral and evolutionary research, especially related to mate choice and species formation. The calls produced by the males are easily replicated and played back under a variety of conditions and crickets are often sedentary enough to be monitored in the field for short periods. They are also easily kept and raised in lab conditions.

An obvious problem with this approach is that it works well for understanding commonalities of species. But it is a drawback in terms of studying how species are different from one another. For example, we might have a good idea of how the genes of a butterfly species control the coloration of the wing. If we see a different species of butterfly with a different wing pattern we probably have a good idea as to what genes are involved but the study of the first species is not going to tell us how those genes are different in the second species.
The new fields of comparative genomics and bioinformatics have at least partially bridged this gap. Automated sequencing technology has allowed the creation of extensive datasets of DNA sequences from many many species. Sequence data from model organisms can be compared to the sequences of many other species to pull out candidate genes for specific functions. So you don't have to start from scratch each time.
Why bring this up now? Well to emphasize a point. We may have a general model of how colors are produced on the surface of an organism. We also know in many cases how those color patterns are inherited and in a few cases like the butterfly wings we know details about the developmental control of color pattern.
In other words we have a good general model of how color pattern would be controlled but for the great majority of the vast array of living things we don't know the details. However we now have much better tools for comparing different species than have existed in the past. This is one reason that biodiversity is so important. It is the way we can expand our biological knowledge. And we can't do that if it isn't there.
II. Color changes
Now to move on to some decidedly non-model organisms. Ones that can change color.
The animals we think of most frequently, birds and mammals, are unusual in having an external covering of dead tissue (feathers or hair). This tissue usually contains pigments or structural colors but because it is dead there is no way to change the color once the hair or feather is formed. Changing color requires replacing feathers or hair. Many birds and mammals change color seasonally which requires loss of existing feathers/hair and growth of new ones. Birds frequently change from breeding to non-breeding plumage for example and many animals living in the arctic switch to a white coat/plumage in the winter.
For animals with coloration in the skin there is the possibility of more rapid color change. Areas of bare skin in mammals and birds are often used in social signaling, presumably because they are more capable of change than fur or feather.
Many reptiles, amphibians, and fishes are capable of fairly short to medium term color change. They have pigments stored inside living cells, either at the body surface or somewhat deeper. Changing the amount of pigment or the location of pigment can affect body color. For example many reptiles turn a very dark color when they are cold to maximize the intake of heat. Other color changes relate to social signaling - the same sorts of changes that happen when birds produce breeding plumage.
Of particular note are species that can produce dramatic and rapid color changes. We'll look at a few examples and consider how it is done.
A couple of local examples in the southeastern US are the squirrel treefrog (Hyla squirella) and the green anole (Anolis carolinensis), sometimes inaccurately called a chameleon. Both of these animals live primarily on vegetation (and nowadays on human structures). They are capable of changing color from bright green to brown. This ability allows them to blend in with both a bark and a leaf background.


Anolis carolinensis, two color phases
The two most dramatic and well known examples of rapid color change are the cephalopods (octopus and relatives) and the true chameleons. In both cases color change occurs through a similar mechanism but the reason for the color changes is quite different.
Chameleons are a group of highly specialized lizards. About half the existing species are found on the island of Madagascar and half on mainland Africa. One species ranges into southern Spain and a few others into southern Asia as far east as India. They are instantly recognizable because of several unique features. Their toes are fused so that each foot has become a clamp, the eyes are encased in cone that can swivel and focus independently, they have immensely long extensible tongues, and the males often have crests and horns reminiscent of those of dinosaurs. Oh, and they can change color two.
How do they do it? The colors are produced by cells known as chromatophores that occur in layers. Some of these cells contain pigments, red and yellow pigments in an upper layer and melanin (dark pigment) in a deep layer. In between is a layer of cells containing guanine cystals which can produce blue color structurally. Anoles and other reptiles and amphibians have similar layers. The difference is largely in the speed and fine control of the changes. When the pigment or crystal is concentrated in the center of the cell it appears transparent. If it is spread out evenly in the cell that color is expressed. By varying the concentration of each cell type the chameleons can rapidly change the color of any part of their body.
If you search the web you will often see it stated that the widespread idea that chameleons change their color to match their background is a myth. Studies of the large chameleon species commonly kept in captivity do show that color change may be more important for social communication and temperature control in these species than for blending in to the background. However, for tiny forest floor living dwarf chameleons color change does seem to be important for camouflage. In fact studies have shown that the nature of the change varies depending on if the chameleon sees a bird (high visual acuity) vs a snake (low visual acuity).
Probably the champion color changers are the cephalopods. Again this is a highly specialized group of animals. One way to think of them is snails that evolved to become fish. They are mollusks, related to snails and clams, but unlike those animals they are highly active with the most highly developed nervous systems of any non-vertebrate.
They are also famous for extremely rapid and complex color changes. All cephalopods except for the chambered nautilus (a very ancient form that diverged from the lineage of other living cephalopods some 470 million years ago) have at least some color changing ability although it is much more pronounced in shallow water species than in open ocean or deep sea species.
The mechanism for producing the colors is similar to that of the animals we have already discussed in that three layers of chromatophores are involved. There are a couple of important differences. One is that some species, particularly deep sea forms, have photophores as well, cells that can generate light and thus influence color from the inside. Most importantly the chromatophore size is influenced by muscles under direct nervous control. So, unlike, the lizard and amphibian examples discussed above, it is direct control by the nervous system rather than hormonal control that drives color change. This means that the changes can be extraordinarily rapid and extremely local. It is possible to see colors move across the surface of the animal.
This video shows light producing organs on the 'vampire squid from hell' about half way through. Attenborough says the light on the tentacles is from bacteria rather than produced directly by the animal. But genuine photophores would look the same and the video was too cool too pass up.
I've included the video below about the mimic octopus although it is more about shape than color because it is one of the most amazing things I've ever seen.
III. Perception of color
This is a bit of a teaser for next time. In the first diary we talked about color as being the wavelengths of light reflected by an object. In a strict sense that is true. But perceived color is also a function of the sensitivity of the eye to wavelengths of light. And not all eyes can see all wavelengths of light. The ability to see colors is a function of the sensitivity of different photoreceptor cells in the eye to different colors. Our eyes are not receptive to either ultraviolet or infrared wavelengths. Surfaces that reflect those wavelengths but not visible light would look black or grey to us but not to organisms that can see those wavelengths.
Our retinas contain two types of photoreceptors, rods and cones. Cones are involved in color vision. Humans have a trichromatic color vision system with three types of cones with pigments sensitive to particular wavelengths. Rods detect light but do not differentiate between colors. They are more sensitive to low levels of light.
Perception of light varies greatly among animals. Insects can see UV light but usually cannot see red light. Many mammals, other than primates, have reduced color vision. Many non-mammalian vertebrates have four or more different types of color detecting receptors in their eyes. For most species the color vision of these animals is not well understood but it seems likely that in many if not most case they can make more subtle color distinctions than we can. Geckos for example, have color vision in extremely low light conditions, when humans are completely color blind. Some invertebrates have highly developed color vision as well. A mantis shrimp apparently has twelve types of color photoreceptors!
So there may be a lot of interesting stuff going on with color that we don't even notice. We'll delve into this in more detail next time when we look at color and plant/animal coevolution.
