In previous diaries, we showed how:
chronic stress mounts central stress networks to increase central stress signaling, which leads to increases in fear and anxiety, hyperarousal and vigilance, addiction, impulsiveness and sensation seeking, long-term changes in memory, and how stress-related juices amplify emotional memory. In this diary we will look at an anatomical neuronal macrosystem, the so-called "extended amygdala," and how different parts of the extended amygdala may be processing cues for specific aspects of aversive experience, namely stone-cold fear versus more "free-floating," or generalized anxiety.
Long-term potentiation
First, let's take another look at the importance of associative emotional memory formation. We previously examined the neural pathways underlying associative fear conditioning in the cortical aspect of the amygdala. We also saw how pairings of an arbitrary stimulus, such as a tone or light, with a biologically significant stimulus, such as a footshock, resulted in the long-term potentiation of neuronal responses to the arbitrary stimulus, how a once innocuous stimulus becomes richly invested with emotional meaning. In fact, "long-term potentiation" is a technical term used to describe the artificial induction of memory traces by electrically stimulating different parts of the brain, such that paired stimulations simulate pairings of conditioned and unconditioned stimuli, and result in long-term increases in responsiveness to formerly innocuous stimuli. The cartoon below depicts one example of how this can be achieved.
In the top panel of this figure, are two neurons, a presynaptic neuron on the left and a post-synaptic neuron on the right. The circular part of each neuron represents the cell body (and dendrites, ie, the antennae), the long process channels signals from left to right, and the little "v" at the end represents the synaptic cleft into which chemicals are released to excite the post-synaptic neuron. First, to get a baseline measure of the post-synaptic neuron's excitability, so we weakly stimulate the pre-synpatic neuron, and then measure the excitatory post-synaptic potential of the post-synaptic neuron.
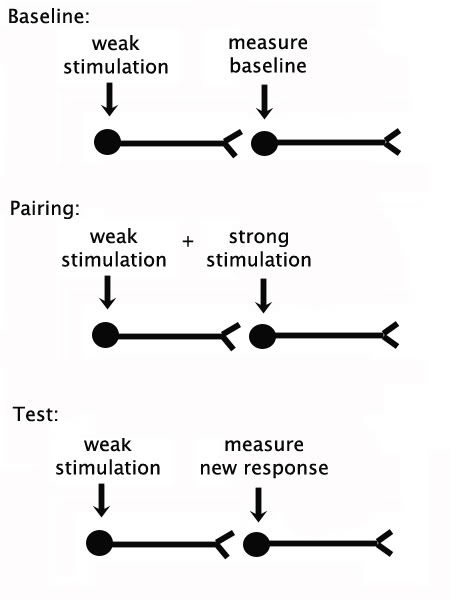
Second, to induce long-term potentiation, weak stimulation of the first neuron is paired with with strong stimulation of the second, as if we were pairing a tone with a shock. As a control, we also want to have the same number of unpaired stimulations to show that the effect is truly associative, that it only occurs when the neurons are stimulated in tandem. So, the unpaired group gets the same number of weak stimulations and strong stimulations, but they never happen at the same time. Finally, to know if this little system of neurons has changed consequent to paired stimulations, we simply repeat Step 1 and weakly stimulate the first neuron, then test the response in the second neuron. Let's take a look.
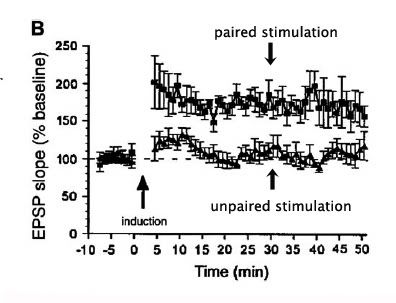
In this image below, we can see time on the horizontal axis, and excitatory post-synaptic potentials (EPSPs) on the vertical axis. The baseline EPSPs are shown to the left the the arrow indicating the induction event (pairing or not pairing the stimulations). Following induction, we can see that the unpaired stimulations do not raise postsynaptic output above baseline, whereas the paired stimulations significantly increase the output of the post-synaptic neuron for a long period of time. This example happens to show long-term potentiation in the cortical aspect of the amygdala (basolateral amygdala). This is precisely what is observed when, say, a formerly neutral stimulus, such as a tone or magazine click is paired with biologically significant aversive events. From now on, that formerly unremarkable stimulus is now richly invested emotional responses. (And what was that great Canadian's seminal hypothesis? Neurons that fire together wire together. Chalk-up a big one for the Canucks and D.O. Hebb!)
The Extended Amygdala
Let's take a look at the anatomy of the brain to see what systems are going to be recruited by such associative events. The image below shows a horizontal section of brain (imagine taking a horizontal knife and cutting on level from your eyeballs to the back of your head. In this picture taken from rats, the eyeballs are above the top of the image, whereas the back of the head is at the bottom).
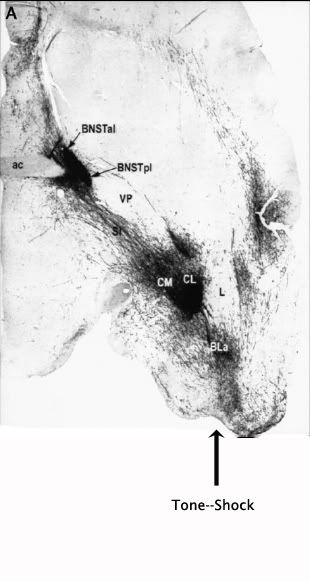
At the very bottom of this picture, I indicated where associations in the cortical aspect of the amygdala (tone--shock, for example) take place. In this picture, the cortical amygdala (BLA) was injected with a tracer that fills neuronal bodies and processes with a goo that can be stained (the black stuff) at a later date to show to where the neurons are projecting. As we saw before, the first principal output nucleus is the central nucleus of the amygdala (CeA; here CL and CM for central-lateral and central-medial, respectively), which ramps up CRF production in response to stress. In addition to this very dense projection, the cortical amygdala projects right through the central nucleus to provide a second very dense projection to the bed nucleus of the stria terminalis (BNST). The BNST also has rich supplies of CRF, a critical anxiety and arousal molecule. This complex of brain nuclei has been referred to as the extended amygdala. Let's look at what types of fear and anxiety-related outputs this complex controls.
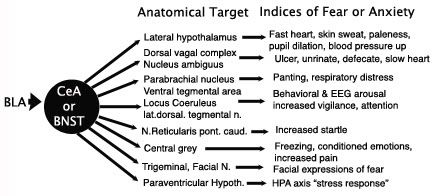
The image above (modified from Walker et al, 2003) depicts our critical structure for emotional associations, the cortical amygdala (BLA) banging smack into the two critical structures of the extended amygdala, the CeA and the BNST, both of which subsequently bang into numerous downstream structures controlling everything from heartrate, breathing, ulcers, startle reactions, and so on, including the the prototypical "stress response" itself. The list is not complete, but you get the idea. If you like to freak yourself at the horror show, or reading Stephen King novels, this is your system. It is also the system that saves your life when danger is real in the here and now, or just around the corner. It is reactive to danger, predicts imminent danger, and can be engaged for generalized mistrust of the environment.
Fear versus Anxiety
A quick word on the difference between fear and anxiety. Fear is the more immediate form of aversive emotion, the type of emotion wherein danger is imminent, where there are specific discrete stimuli, such as a tone, a magazine click, or a window breaking, indicating that trouble is "here" now, and which initiates fight or flight responses, such as freezing, biting, running, and jumping. Anxiety is a more diffuse form of emotion that is not indicated by or pinned on to specific discrete stimuli, but rather by more general places or contexts or "airs" that don't indicate that trouble is here now, but is perhaps around some corner, down the road, or may be calling soon.
Enhanced Startle Responses
A number of studies have investigated which specific areas of the brain mediate the effects of various classes of external and internal cues on fear and anxiety responses. In the "startle response" paradigm, rats are simply exposed to a sudden noise burst and the amplitude of their startle response is measured, e.g., how much does the floor shake as a result of their startle reaction.
In the graph below (adapted from Lee & Davis,1997), rats were first subjected to a noise burst to measure their baseline startle responses. Then they were exposed to either an endogenous anxiety cue, in this case CRF, which induces generalized anxiety (left panel), or a discrete exogenous cue (I believe it was a tone) that was formerly paired with a footshock (right panel). There were 3 groups of rats: Intact controls, and rats bearing lesions of either the central nucleus of the amygdala (CeA) or the bed nucleus (BNST). In the presence of CRF-induced generalized anxiety, the intact animals showed a large increase in startle amplitude, as expected. Lesions of the CeA did not significantly impair this response, whereas lesions of the BNST essentially blocked the effects of anxiety induction. However, when the tone associated with footshock is presented, lesions of the CeA blocked the increase in startle, whereas lesions of the BNST had little effect.
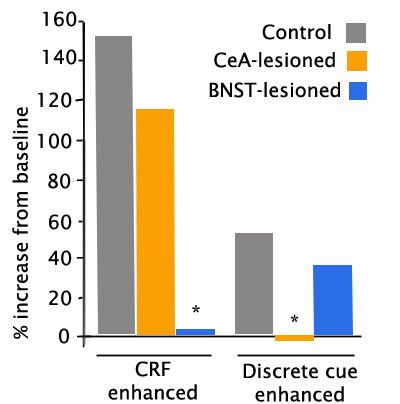
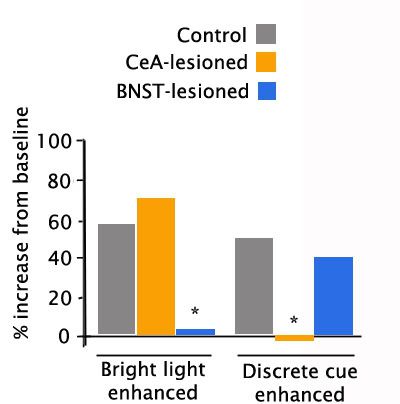
A similar study (Walker & Davis, 1997) compared a different generalized anxiety cue (bright light, which rats dislike) to a discrete cue formerly associated with shock. Once again, the enhancement of the startle response to the noise burst given in the presence of the generalized anxiety cue that contained no specific warning was unaffected by CeA lesions, but was virtually abolished by BNST lesions (left panel). And once again, when a specific threat was presented (a tone predicting footshock, with no actual shock given), the enhanced startle was unaffected by BNST lesions, but was abolished by lesions of the CeA. These studies strongly suggest that CeA mediates specific immediate threats (red alerts), whereas BNST mediates a more generalized anxiety when specific information is unavailable (orange alerts).
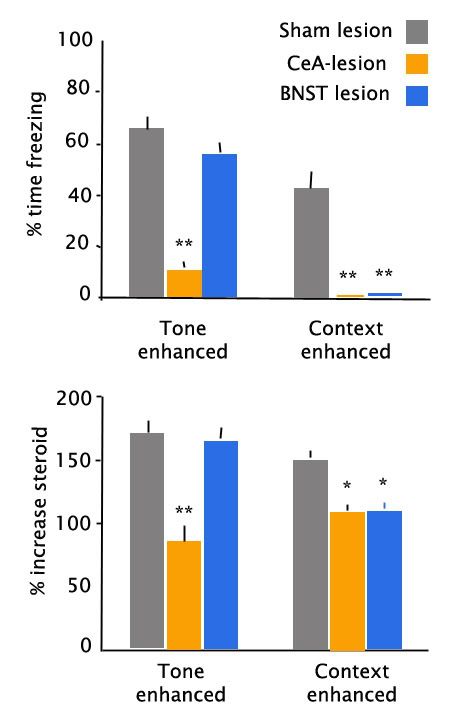
These separable functions within the extended amygdala are not perfectly exclusive, and for more intermediate predictors of specific threat, they seem to have some overlapping function. In the graph below (adapted from Sullivan et al, 2004) the authors measured the enhancement of freezing behavior and stress steroid secretion in the presence of either a discrete stimulus (tone) predicting footshock or a context predicting footshock. As before, when there was a specific discrete threat (top left panel), CeA lesions nearly abolished the enhancement in freezing behavior, whereas BNST lesions had no effect. However, in the presence of a slightly more general threat, both CeA and BNST lesions abolished the cue-enhanced freezing. Similar findings held for cue-induced increases in stress steroid secretion (bottom panels). Thus, the extended amygdala appears to mediate control over a continuum of threat assessment.
Behavior Systems
I found it interesting when reading through some of the clinical lit on PTSD that someone had applied a "Behavior Systems" view as an explanation of PTSD. It would take way too long to explain this adequately, so I'll just give you a quick snapshot. Keep in mind, this is highly speculative to begin with, but conceptually interesting. In the Behavior Systems view of animal behavior, animals are functionally organized into major modules according to major functions, e.g., feeding, mating, reproduction, socializing, defense/aggression, and so on. These major, species-specific functions are subdivided into hierarchies of pre-organized lesser modules that "funnel" animals into ever-closer approximations of solutions for specific problems. For example, in the feeding system in rats, increasing steroid levels near the time of waking facilitate a major bout of locomotor activity, presumably related to the problem of finding breakfast. This module of "general search" includes path following behaviors, thigmotaxis (wall hugging), whisking and smelling, and so forth, i.e., it is a set of behavioral and perceptual dispositions designed to put the animal in contact with more specific stimuli related to food. Once the animal locks on to more specific cues, such as an odor trail, it advances into a more "focal search" routine involving handling and sampling, or if preying on animals perhaps chasing or lying in wait for prey. As cues become more specific, appropriate sub-routines are released that are relevant to capturing, killing, ingesting, and so on. If the animal fails, it progressively backs out of the more specific routines into more general routines to put itself back on track of the appropriate cue-level. In addition, each of these modules can be conditioned to various stimuli that are noticeable during each routine. Thus learned cues can evoke different phases of these routines.
In the defensive system, a continuum of "predatory imminence" includes various temporal and spatial distances from attack, and several distinct phases have been identified, including pre-encounter, post-encounter, circa-strike, and recuperative phases. In the pre-encounter phase, before a predator has been identified, animals may organize their other modules to minimize the chance of predatory encounters. For example, a prior history of predatory encounters will cause rats to re-organize their foraging bouts to include fewer bouts leaving the nest to forage and take larger meals each time they feed to defend body weight. Once a predator is identified or suspected (a twig snaps!) in the post encounter phase, two behaviors are dominant: fleeing if possible and highly likely to succeed, or more likely freezing, as inanimate objects are much more difficult to detect. When predatory imminence increases at around the time of the strike, then running, jumping, biting, screaming, and stress-induced analgesia predominate. In the post-strike recuperative phase, hypoalgesia promotes attention to wounds, while depressive-like hypoarousal promotes rest and avoidance of further encounters.
To make a long story short, the author argued that various aspects of this continuum of predatory imminence in the defensive system become excessively amplified or dysregulated in PTSD, thus explaining various symptoms and dissociative states. As I said, this is involved and speculative.
However, based on some of the data we just looked at, it certainly appears that different parts of the extended amygdala are mediating different behaviors and other motor outputs depending upon the specificity of the danger cues, as if they existed on different domains of the continuum of predatory imminence, with the central nucleus of the amygdala predominately processing cues closer to the circa-strike end of the continuum, i.e., cues indicating naked fear, and BNST processing cues closer to the pre-encounter end of the continuum, i.e., cues indicating generalized anxiety in the absence of specific cues.
